 FIGURE 5.1 Gate control theory as originally schematically described by Melzack and Wall2 where large-diameter (L) and small-diameter (S) primary afferent fibers project to substantia gelatinosa (SG) and second-order transmission (T) neurons in the spinal dorsal horn. The inhibitory effect of SG neuronal activity is increased by L and decreased by S fiber activity. T neurons transmit information to the brain and other action sites. Activation of peripheral or central projections of L fibers using transcutaneous nerve stimulation, peripheral nerve stimulators, or dorsal column stimulators would all be expected to produce inhibition of S fiber input to the T cells. |
 FIGURE 5.2 Wind-up responses of single dorsolateral column axon to repeated stimulation of the sural nerve at sufficient intensity to activate A and C fibers (no wind-up seen with A-fiber stimulation by itself). The vertical time markers on the far right represent 100 milliseconds. Each mark at the bottom of the time line represents the stimulation artifact and the burst of activity immediately above each of these stimulations is the response to A-fiber stimulation (each dot represents an action potential). The more delayed responses are to the more slowly conducting C-fiber inputs. Response to stimulation shows increasing C-fiber wind-up responses on to 1 per second stimulation (not to 1 every 2 or 1 every 4 second stimulation rates at right). Wind-up lasts for only several seconds following the stimulation as seen by transient increase in spontaneous activity. (Redrawn from Mendell LM. Physiological properties of unmyelinated fiber projections to the spinal cord. Exp Neurol 1966;16:316-332.) |
lability can occur. The precise neurophysiologic circuits associated with this descending inhibition is of significant debate, but known inhibitory neurotransmitters such as norepinephrine (NE) and serotonin (5-hydroxy-tryptophan; 5-HT) are synthesized in the brainstem and transported to the spinal cord from multiple supraspinal sites. This role for supraspinal structures in providing descending influences on spinal reflexes has long been recognized. In 1915, Sherrington and Sowton12 demonstrated enhanced flexion reflexes following spinal transection. Later in 1926, Fulton13 suggested that this effect reflected removal of tonic descending inhibitory modulation of spinal interneurons mediating those reflexes. Descending control of flexion reflexes was extensively studied in ensuing years,14 but these studies did not target the issue of how the brain might specifically modulate incoming nociceptive signals from peripheral tissue.
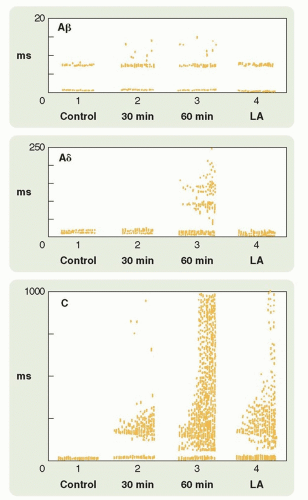 FIGURE 5.3 Raster dot displays of a single biceps femoris unit activated by stimulation of the sural nerve once every 2 seconds before an ipsilateral thermal injury (Control), 30 and 60 minutes postinjury, and 10 minutes after the injured foot has been completely anesthetized with local anesthetic (LA). Each dot represents a unit discharge. The vertical scale is the latency of the responses after sural nerve stimulation, and the stimulus artifact can be seen at time 0. Stimulation strengths were sufficient to activate Aδ, Aδ, and C fibers. Note the different time scales used in the three panels to record the activity evoked by the three different fiber populations. In the preinjury state, only Aδ input was evoked. Thirty minutes after injury, a C-fiber response begins to occur, whereas at 60 minutes, both Aδ and C-fiber evoked responses are present (the C-fiber responses with wind-up). Ten minutes after LA, the C-fiber evoked responses remain higher than before the injury suggesting a central component of the sensitization. (Redrawn from Woolf CJ. Evidence for a central component of post-injury pain hypersensitivity. Nature 1983;306:686-688.) |
 FIGURE 5.4 A modification of the gate control theory schematic models includes excitatory (white circle) and inhibitory (black circle) links from the substantia gelatinosa (SG) to the transmission (T) cells as well as descending inhibitory control from brainstem systems. The round knob at the end of the inhibitory link indicates that its actions may be presynaptic, postsynaptic, or both. All connections are excitatory except the inhibitory link from SG to T cells. (Redrawn after Melzack R, Wall PD. The Challenge of Pain. New York: Basic Books; 1983.) |
midbrain reticular formation, locus coeruleus/A6 cell group (LC/A6), the A5 cell group, the lateral reticular nucleus (LRN) and nearby A1 and C1 cell groups, the parabrachial nucleus/A7 cell group (PBN/A7) and the nucleus tractus solitarius (NTS). A limited amount of information is also available on cortical and limbic systems such as the anterior cingulate cortex (ACC), the amygdala, and hypothalamus that contribute to descending modulation. It is the investigation of these structures that has led to our current understanding of how descending pain modulatory systems affect pain perception. This topic has been well reviewed.1,23,24,25,26,27,28,29 Table 5.1 summarizes results related to many of these areas separately along with the neurotransmitters associated with their putative inhibitory versus facilitatory effects on nociceptive transmission. A summary of the most important components is described in Figure 5.5.
TABLE 5.1 Central Nervous System (CNS) Sites Modulating Nociceptive Transmission (NT) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Related posts:




Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


