CHAPTER 6 Thermoregulation: Physiology and Perioperative Disturbances
One of the many physiologic adaptations required for the survival of homeothermic species is the ability to maintain constant core body temperature within narrow limits. The significance of thermal regulation for neonates was first appreciated by two French obstetricians, Tarnier and later Budin, who in 1907 published the finding of significantly higher survival rates in normothermic versus hypothermic neonates (Budin, 1907). Several other investigators later confirmed the importance of thermal stability in the adaptive process and further elucidated the mechanisms by which neonates, infants, and children are able to behave as homeotherms (Silverman and Blanc, 1957; Cross et al., 1958; Silverman et al., 1958; Bruck, 1961). A homeothermic organism is characterized by its ability to maintain constant core (or central) body temperature despite changes in the ambient temperature. Not many physiologic parameters are as vigorously and effectively controlled as the core temperature. In humans, the central body temperature refers to the temperature of the vessel-rich group organs (i.e., brain, heart, lungs, liver, and kidneys) and is normally maintained within ±0.2° C of its set point of 37.0° C. This so-called interthreshold range defines the limits within which no thermoregulatory effector responses are triggered and the human organism behaves poikilothermically. The musculoskeletal system makes up the major part of the peripheral compartment, which is considered to be a dynamic buffer in the thermoregulatory system. The skin, representing the shell compartment, acts as a barrier to the environment.
Temperature control is subjected to circadian rhythms, some of which begin in the first days of life. The circadian rhythm of body temperature is generally age dependent (less pronounced at very young and very old ages) and generally closely associated with the sleep-wake cycle. In fertile women, a monthly rhythm in body temperature exists because of a higher set-point temperature in the luteal phase of the menstrual cycle (Hardy, 1961). Despite the fluctuations of body temperature within these rhythms, temperature control is tight and is accomplished by a sophisticated system that balances heat production and heat loss.
Accidental hypothermia occurs commonly in patients of any age who are undergoing anesthesia and surgery, and it has often been accepted as an unfortunate but unavoidable consequence of the surgical procedure. This high rate of hypothermia led Pickering to his famous statement, “The practical difficulty in cooling men is to break through the defenses of the body; the most effective means is to give an anesthetic…” (Pickering, 1958).
Temperature monitoring
Perioperative detection of changes in body temperature requires appropriate monitoring and monitoring sites. Most national anesthetic societies now have guidelines that require that one method for measuring body temperature during anesthesia be available (La Société Française d’Anesthésie et de Réanimation, 1994; American Society of Anesthesiologists, 2005; Australian and New Zealand College of Anaesthetists, 2006; Canadian Anesthesiologists’ Society, 2007).
Infrared thermometers (thermopiles) are quite popular in postanesthesia care units and on hospital wards; however, for continuous temperature monitoring during anesthesia they are not suitable. Despite a fast response time, their accuracy in clinical practice has not been confirmed (particularly when they are not used properly) (Craig et al., 2002; Heusch and McCarthy, 2005; Leon et al., 2005; Dodd et al., 2006).
Temperature-sensitive liquid crystals have been used to measure skin temperature. Although these devices are easy and convenient to handle, they generally do not meet the accuracy criteria required for clinical use. Their readouts can easily be affected by changes that are related not only to body temperature but also to skin blood flow (Leon et al., 1990; MacKenzie and Asbury, 1994). The suggestion of simply adding a constant correcting value (e.g., 2.2° C) to an arbitrary skin temperature to estimate central temperature has been shown to be unreliable (Burgess et al., 1978; Leon et al., 1990).
Body temperature varies widely within the body. Because of their high perfusion rates, core tissues tend to maintain a constant temperature, whereas peripheral tissues usually have significantly lower and less uniform temperatures that may differ by several degrees within a short distance from each other (Colin et al., 1971).
It has been suggested that hypothalamic temperature reflects core temperature, although there is no physiologic evidence that hypothalamic temperature precisely represents central temperature (Benzinger, 1969). Core-temperature measuring sites recommended for clinical use are the tympanic membrane, nasopharynx, distal esophagus, pulmonary artery, and, with some limitations, bladder and rectum. These sites usually provide equal readings in humans who are awake and in those who are anesthetized and undergoing noncardiac surgery (Cork et al., 1983). However, different temperatures may be measured at different monitoring sites and under certain clinical conditions, and the physiologic and clinical significance of these differences may vary.
The precision and accuracy of measurements at different body sites have been studied, and each site has its advantages and disadvantages (Cork et al., 1983; Bissonnette et al., 1989b). Ideally, the temperature-monitoring site reflects core temperature and is associated with only minimal or no morbidity.
Skin temperature measurements offer little as a reflection of core temperature (Lacoumenta and Hall, 1984; Bissonnette et al., 1989b). Because there is a wide variation in skin temperature, depending on the site of monitoring, several investigators have suggested monitoring between 4 and 15 sites, using different formulas to accurately describe the mean skin temperature (Shanks, 1975; Puhakka et al., 1994; Ram et al., 2002). For skin temperature to be of clinical value, it must closely reflect central temperature in the perioperative setting so that mild hypothermia and early signs of malignant hyperthermia (MH) can be detected. Beside the fact that increased body temperature is a late sign of MH, it is unlikely that skin temperature correlates well with central temperature during the early stages of MH, because circulating catecholamine concentrations may be up to 20 times higher than normal and result in significant changes of skin perfusion (Sessler, 1986; Sessler and Moayeri, 1990).
Tympanic-membrane temperature has been suggested as the most ideal temperature-monitoring site. Although it is not necessary for the temperature probe to be in direct contact with the tympanic membrane to accurately reflect tympanic temperature, the external auditory canal needs to be sealed by the probe to allow the air column trapped between the probe and the tympanic membrane to reach a steady-state temperature. During the initial postoperative period after infants and children have had cardiac reconstructive surgery, tympanic temperature does not correlate well with brain temperature and therefore does not provide a reliable estimate of central body temperature (Muma et al., 1991; Bissonnette et al., 2000). Because of difficulties associated with obtaining appropriate-sized thermistors and reports of tympanic membrane perforation, the clinical use of continuous intraoperative temperature measurement has been discouraged.
In contrast, oral temperature is generally considered less adequate and is therefore not recommended as an accurate site for intraoperative temperature-monitoring (Cork et al., 1983).
Esophageal temperature probes are often combined with an esophageal stethoscope, which makes this site particularly attractive for the pediatric population. In infants and children, and in patients who are cachetic, the thermal insulation between the tracheobronchial tree and the esophagus is minimal. Therefore, the respiratory gas flow may result in erroneous temperature readings, particularly when the fresh gas flow is high and its temperature differs significantly from body temperature (Bissonnette et al., 1989b). Furthermore, central temperature is measured only if the tip of the probe is placed in the distal third of the esophagus at the point where the heart sounds are the loudest (Bissonnette et al., 1989b; Stoen and Sessler, 1990). In patients with endotracheal tubes, monitoring of esophageal temperature is more reliable than rectal temperature and more practical than tympanic temperature.
Axillary temperature is not only the most commonly used method of measuring temperature, but it is also the most convenient site for temperature monitoring. It has been reported to be as accurate in measuring central temperature as tympanic membrane, esophageal, and rectal temperature sites. However, this accuracy is only achieved when the tip of the thermometer is carefully placed over the axillary artery and the arm is closely adducted (Bissonnette et al., 1989b). Unfortunately, malpositioning of the probe may result in unreliable estimates of core temperature. Infusion of cool solutions at high flow rates in small children on the ipsilateral side of the thermometer probe may result in falsely low temperature readings.
Rectal temperature monitoring can provide a central temperature reading; it is associated with minimal morbidity and its ease of insertion confers major advantages (Bissonnette et al., 1989b). Problems to be considered with its use pertain to the probe’s insulation by feces, its exposure to cooler blood returning from the legs, the influence of an open abdominal cavity during laparotomy, or irrigations of the bladder or the abdomen with either cold or warm solutions. Relative contraindications for rectal temperature probe insertion are inflammatory bowel disease, neutropenia and/or thrombocytopenia, and the need to irrigate the bowel or bladder.
Bladder-temperature monitoring is considered to be one of the most accurate methods of measuring core temperature. Its precision has been demonstrated to be identical to pulmonary-artery–temperature monitoring as long as urinary output is high; however, when urinary output is normal or less than normal, this site may become inaccurate in reflecting central temperature (Horrow and Rosenberg, 1988; Brauer et al., 2000).
Physiology of thermal regulation
Survival from body temperatures as low as 13.7° C has been reported, whereas death resulting from protein denaturation occurs within 7° C above normality at approximately 44° C (Gilbert et al., 2000). This illustrates a tolerance for cold that is more than three times higher than that for heat, which explains why the system for heat dissipation needs to be much more effective than the system for defense against cold.
The thermoregulatory system is similar to other physiologic control systems in the sense that the brain uses negative feedback mechanisms to keep temperature variations from normal values minimal. The principal site of temperature regulation is the hypothalamus, which integrates afferent signals from temperature-sensitive cells found in most tissues, including other parts of the brain, spinal cord, central core tissues, respiratory tract, gastrointestinal tract, and the skin’s surface. The processing and regulation of thermoregulatory information occurs in three stages: afferent thermal sensing, central regulation, and efferent response (Fig. 6-1).
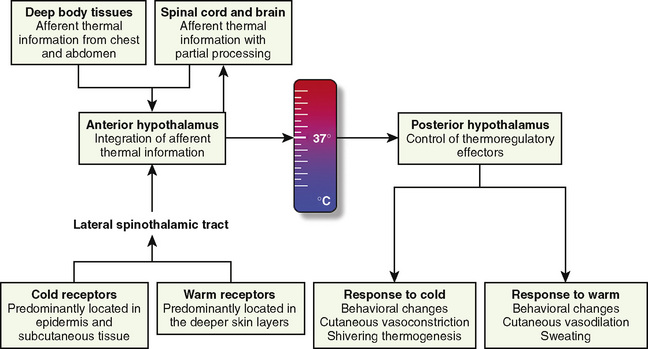
Afferent Thermal Sensing
Anatomically distinct warm and cold receptors in the body periphery sense the ambient temperature. The skin contains about 10 times more cold receptors than warm receptors, acknowledging the important function of the skin in the detection of cold (Poulos, 1981). Thermosensitive receptors are also located in close proximity to the great vessels, the viscera, and the abdominal wall, as well as in the brain and in the spinal cord. Each receptor type transmits its information through an afferent-nerve–conduction pathway. Although information originates from anatomically different nerve fibers, the speed of transmission is mainly influenced by the intensity of the stimulus rather than the type of nerve fiber. It is well established that the rate of change in skin temperature alters the apparent importance of the change. Rapid changes contribute up to five times as much to the central regulatory system as slower changes with comparable intensity (Wyss et al., 1975). However, other than in patients undergoing cardiopulmonary bypass surgery (where rapid temperature changes are common), the rate of change in core temperature does not appear to substantially influence the magnitude of the provoked regulatory responses.
Thermal information gathered from peripheral warm receptors, whose maximal discharge rate is between 45° and 50° C, is carried by unmyelinated C fibers. These C fibers also convey pain sensations, which explains why intense heat cannot be distinguished from severe pain (Pierau and Wurster, 1981; Poulos, 1981). Although most ascending thermal information travels along the spinothalamic tracts in the anterolateral spinal cord, no single spinal tract is solely responsible for conveying thermal information (Hellon, 1981).
Central Regulation
The preoptic area of the hypothalamus contains cold- and heat-sensitive neurons, with the latter predominating by a ratio of 4:1 (Boulant and Bignall, 1973). However, the vast majority of the neurons in this area are insensitive to temperature (Nakayama et al., 1963). This area also receives and processes nonthermic afferent information, which seems to be important in controlling the adaptive mechanisms and the behavior of the organism (Hori and Katafuchi, 1998).
Direct heat stimulation of this area results in increased discharge rates from the heat-sensitive neurons and activation of heat-loss mechanisms. Conversely, hypothalamic cold-sensitive neurons respond with increased discharge rates to direct cooling of the preoptic area of the hypothalamus (Boulant, 1974; Boulant and Demieville, 1977). Other centers involved in thermoregulation include the dorsomedial hypothalamus, periaqueductal gray matter, the nucleus raphe pallidus in the medulla oblongata, and the spinal cord, although their functions are not yet fully elucidated (Guieu and Hardy, 1970; Simon, 1974; Cabanac, 1975; Dickenson, 1977).
The contribution of the central thermoreceptors to thermal regulation under normal conditions is limited by the marked predominance of thermal input from peripheral receptors (Downey et al., 1964). These central receptors take over thermoregulation if the sensory input from peripheral sensors is disrupted (e.g., through central neuraxial anesthesia or spinal cord transection), but they are less efficient when compared with peripheral thermoreceptors (Downey et al., 1967).
The threshold temperature defines the central temperature at which a particular regulatory effector is activated (Box 6-1). When the integrated input from all sources is signaling that the interthreshold range is exceeded on either side, efferent responses are initiated from the hypothalamus to maintain normal body temperature.
Box 6-1 Definition of Temperature Regulation Terms
Efferent Response
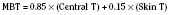
where T denotes the temperature measured in °C. Other formulas do exist (Ramanathan, 1964; Colin et al., 1971; Shanks, 1975; Puhakka et al., 1994). See the following example:
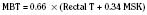
where MSK reflects the mean skin temperature (in °C), which then equals:
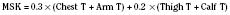
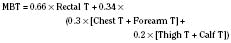
Skin temperature is the most important parameter in triggering behavioral changes; however, in terms of impact on the thermoregulatory autonomic response, the thermal input from the skin contributes only about 20% (Cheung and Mekjavic, 1995; Lenhardt et al., 1999). The main part of this autonomic response depends on afferent information from the central core, which includes the brain (parts other than the hypothalamus), the spinal cord, and deep abdominal and thoracic tissues, with each of them contributing about 20% to the central thermoregulatory control (Jessen and Mayer, 1971; Simon, 1974; Mercer and Jessen, 1978; Jessen et al., 1984).
The most commonly described thermoregulatory model is a set-point system in which hypothalamic integration of thermal information indicates a body temperature above or below a predetermined threshold and then triggers warm or cold defenses. This set-point model, borrowed from engineering models, provides an easy way to explain how the thermoregulatory system functions and how temperature is regulated. In this model, the body compares its actual central temperature against a set reference temperature and then balances heat loss and heat-generating mechanisms to keep the temperature at this set reference point. However, while convenient, this model may not be accurate. More recent research suggests that peripheral and central thermoreceptors are connected through several other neurons to a thermoregulatory effector cell to form a thermoeffector loop (Kobayashi, 1989). In this set-up, once the temperature reaches the range for which the particular thermosensitive neuron has its highest sensitivity, its firing rate increases significantly and—independent of a central nervous system controller—triggers a response in the thermoregulatory effector. Central body temperature in this model is then the averaged result of all the thermoeffector loop actions combined, basically making a central controller (i.e., the hypothalamus) redundant (Kobayashi, 1989; Romanovsky, 2004). Although this more recent model has received a fair amount of attention, it has not yet been widely accepted in clinical practice.
Regardless of the actual model used, if an effective thermoregulatory system is in place, behavioral responses (e.g., heating the home, looking for shelter, or putting on a jacket) to environmental temperatures outside the thermoneutral range (approximately 28° C for an unclothed adult) remain the quantitatively most important thermoregulatory effectors in humans and are far more efficient than all of the autonomic responses combined. Cutaneous vasoconstriction is the first and most consistent thermoregulatory response to hypothermia. Total digital skin blood flow can be divided into nutritional (capillaries) and thermoregulatory (arteriovenous shunts) components. Cold-mediated decreases in cutaneous blood flow are most pronounced (down to 1% of the normal blood flow seen in a thermoneutral environment) in arteriovenous shunts of the hands, feet, ears, lips, and nose (Grant and Bland, 1931; Hillman et al., 1982). These shunts are typically 100 µm in diameter, which means that one can divert 10,000 times as much blood as a capillary with a 10 µm diameter under otherwise unchanged conditions (i.e., same length and pressure gradient) (Hales, 1985).
Flow changes not only in the arteriovenous shunts, but also in the far more numerous capillaries (Coffman and Cohen, 1971). The impressive decrease in cutaneous perfusion secondary to thermoregulatory vasoconstriction results in a heat-loss reduction of 50% from the hands and feet, but only of 17% from the trunk, resulting in an overall heat-loss reduction of only 25% (Sessler et al., 1991).
Thermal regulation in the newborn
Premature infants, infants who are small for gestational age, and even full-term neonates have an exceptionally large skin surface area compared with their body mass (assuming a normal ratio for a full-term neonate of 1, then the ratio for an adult is approximately 0.40). Heat loss is further increased because there is only a thin layer of subcutaneous fat and reduced keratin content of the infant’s skin, which results in increased thermal conductance and increased evaporative heat loss. (Therefore, when compared with adults, neonates lose proportionately more heat through their skin in similar environments). In contrast to adults, the capabilities and the functional range of the neonate’s thermoregulatory system are significantly limited and easily overwhelmed by environmental factors. The lower ambient temperature limit of thermal regulation in adults is 0° C, whereas that in newborns is 22° C. The combination of increased heat loss and a diminished efficacy of the thermoregulatory response with a reduced ability to generate heat puts these infants at high risk for hypothermia. The same anatomic properties that are responsible for the increased risk of hypothermia also allow for rewarming that is three to four times faster in infants and children compared with adults (Szmuk et al., 2001).
The neutral temperature (or the thermoneutral zone) is defined as the ambient temperature range, at which the oxygen demand (as a reflection of metabolic heat production) is minimal and temperature regulation is achieved through nonevaporative physical processes only (i.e., vasoconstriction or vasodilation). The upper limit of this range is called the upper critical temperature and marks the ambient temperature at which evaporative heat losses are triggered. Similarly, the lower critical temperature defines the ambient temperature below which metabolic heat generation is activated (nonshivering and/or shivering thermogenesis). Depending on the neonate’s weight, this neutral temperature zone is in the range of 32° to 35° C (unclothed in a draft-free environment with uniform temperature and moderate humidity), whereas for an unclothed adult it is approximately 28° C (Hey, 1975). In a thermoneutral environment, the cutaneous arteriovenous shunts are open and skin blood flow is maximal.
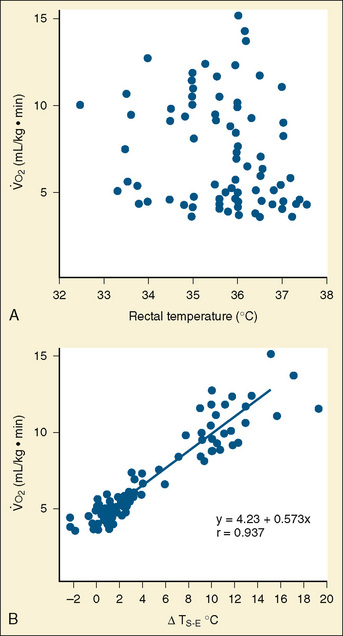
Maintaining core temperature in a cool environment results in increased oxygen consumption and potentially metabolic acidosis. It was demonstrated long ago that oxygen consumption does not correlate with rectal temperature in full-term neonates, but rather it increases directly with the skin surface-to-environment temperature gradient (Adamson et al., 1965). Oxygen consumption is minimal at gradients of 2° to 4° C. Thus, at environmental temperatures of 32° to 34° C and an abdominal skin temperature of 36° C, the resting newborn infant is in a state of minimal oxygen consumption (i.e., in a neutral thermal state). Normal rectal temperature, therefore, does not necessarily imply a state of minimal oxygen consumption in this age group, because the baby could activate all its physiologic defense mechanisms to maintain normal rectal temperature (Fig. 6-2).
Of particular concern in view of thermoregulation in the newborn is the head, which comprises up to 20% of the skin’s total surface area and shows the highest regional heat flux ability (Anttonen et al., 1995). In neonates and infants, the head may account for up to 85% of body-heat losses, which can be explained by the thin skull bones and the usually sparse scalp hair in combination with the close proximity to the highly perfused brain (core temperature) (Fleming et al., 1992). Facial cooling may increase oxygen requirements in the term and preterm infant by up to 23% and 36%, respectively, thereby further demonstrating the effectiveness of protecting the infant from heat loss by covering the head (Sinclair, 1972).
Thermoregulatory vasoconstriction and vasodilation are already present during the first day of life in both the premature and the full-term infant (Bruck, 1961; Lyons et al., 1996). With vasoconstriction, cutaneous blood flow decreases and the effect of tissue insulation increases, which results in an overall reduction in conductive and convective heat losses.
Heat loss mechanisms
The abilities to produce and dissipate heat are fundamental for a homeothermic organism. Controlled heat loss in homeotherms is accomplished in two stages, both governed by the physical laws of conduction, radiation, convection, and evaporation (Box 6-2) (Swyer, 1973). The second law of thermodynamics states that heat can be transferred from a warmer to a cooler object but never from a cooler to a warmer object. What this means is that the warmer object (in the operating room setting, this is almost exclusively the patient) is used to warm up the surrounding cooler objects (e.g., the operating room walls, tables, and instruments). Although most anesthesiologists consider heat loss to be a nuisance, without any heat loss to the environment (i.e., perfect insulation), the body of an awake adult at rest (assuming a metabolic rate of 75 W) would warm up by at least 1° C per hour. (Keep in mind that during exercise, metabolic heat generation can increase up to tenfold.) This can be calculated with the following formula:
Box 6-2 Mechanisms (by Percent) of Heat Loss for a Neonate in a Thermoneutral Environment
| Radiation | 39% |
| Convection | 34% |
| Evaporation | 24% |
| Conduction | 3% |
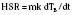
where HSR is the heat storage rate (W), m is the body mass (kg), k is the specific heat coefficient of the human body (3.5 • 103 J/°C), dTB is the change in body temperature (°C), and dt is the time interval (sec) (Burton, 1935).
The first stage of heat loss during anesthesia occurs with the transfer of heat from the body core (central compartment) to the periphery and the skin’s surface, which is referred to as the concept of internal redistribution of heat. In the second stage, heat is dissipated from the skin’s surface to the environment (see “Anesthesia and Hypothermia,” p. 168). Physiologic manipulations of regional blood flow and changes in the thermal conductance properties of the insulation tissue can influence both gradients. Most studies of thermal regulation in infants and children have quantified the relative contributions of radiation, convection, evaporation, and conduction to heat loss. A study in newborns in a thermoneutral environment found radiation, convection, evaporation, and conduction to account for 39%, 34%, 24%, and 3%, respectively, of total heat loss (Hey, 1973). However, the conditions in the operating room rarely meet the criteria for thermoneutrality and the relative contributions of each of these four physical components to total heat loss can vary significantly. Figure 6-3 gives an overview of the heat loss mechanisms involved in the operating room setting.
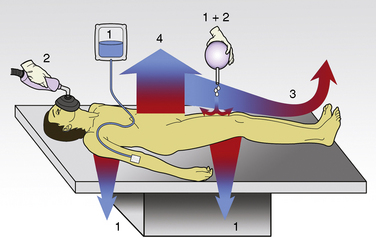
Conduction
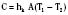
The coefficient hk is a property of the material or interface between the two objects that determines the rate of heat transfer per unit area per unit temperature difference (W/m2 • °C). During surgery, relatively little heat should be lost to the environment via conduction, because the patient is supposed to be well insulated from surrounding objects (Allen, 1987). However, conduction is also responsible for heat loss created by warming up cool intravenous fluids and irrigation solutions, which have the potential to significantly and quickly reduce body temperature. Attention should also be paid to ensure that the patient’s skin is not in contact with any metallic surfaces, because metals have a high thermal conductivity, thereby facilitating heat transfer. (In addition, contact with metallic surfaces during surgery must be avoided to prevent skin burns from electrocautery.) The physiologic factors controlling conductive heat loss are cutaneous blood flow and the thickness of the subcutaneous tissue (insulation).
Radiation
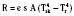
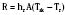
where hr denotes the radiation coefficient, an integration of emissivity and the Stefan Boltzmann constant. Heat transfer by radiation principally depends on the temperatures of the two surfaces concerned and is unaffected by air movement or the distance between the surfaces, and it can take place even across a vacuum (Allen, 1987).
As previously stated, newborns and infants have a large surface area-to-mass ratio, thus radiant heat loss is proportionally greater the smaller the infant. In both the infant who is awake and the infant who is anesthetized, radiation is the major factor for heat loss under normal conditions. The human body is an excellent emitter of energy at wavelengths relevant to heat transfer, and the probability of photon reflection in the standard operating room is almost zero. Radiant heat loss in the operating room is therefore a function of the temperature difference between the patient’s body and the room (i.e., the floor, walls, and ceiling) and all the objects in it. Warming up the operating room (and its contents) reduces not only the temperature gradient between the patient and the environment but also radiant heat losses. However, as long as a temperature gradient exists, the patient continues to warm up the surrounding environment. At a room temperature of 22° C, about 70% of the total heat loss is a result of radiation (Hardy et al., 1941). A simple single-layer covering of the body dramatically reduces the heat loss by convection and radiation; thus, a thin shirt (e.g., a silk blouse, although it provides only negligible insulation) already results in considerably increased thermal comfort.
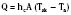
Evaporation
Evaporative heat loss occurs through the skin and the respiratory system. Under conditions of thermal neutrality, evaporation accounts for 10% to 25% of heat loss. Physical factors governing evaporative heat loss include relative humidity of the ambient air, velocity of airflow, and lung minute ventilation. The driving force behind evaporation is the vapor pressure difference between the body surface and the environment. Evaporative losses include mainly three components: sweat (sensible water loss); insensible water loss from the skin, respiratory tract, and open surgical wounds; and evaporation of liquids applied to the skin, such as antibacterial solutions. The evaporation of water from a surface is dependent on energy that is absorbed from the surface during the transition from a liquid to a gaseous state. This energy is called the latent heat of vaporization, and in the case of sweat, it has a value of 2.5 × 106 J/kg. This figure emphasizes the extraordinary power of the human sweating mechanism as a means of dissipating heat, especially considering that an adult in excellent physical condition can produce up to 2 or 3 liters of sweat per hour (Armstrong et al., 1986; Godek et al., 2008). In an environment where the air temperature is equal to or higher than the skin temperature, sweating is the only mechanism available for dissipation of heat that originates from metabolic production. In this situation, anything that limits evaporation, such as high ambient humidity or impermeable clothing, may easily lead to heat storage and a potentially fatal rise in body temperature. Evaporative heat loss can be calculated as follows:
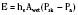
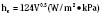
where V is the airflow velocity (m/sec). The important point to note is that evaporation is determined by the vapor pressure gradient between the exposed body surface and the ambient air and the rate of airflow across the surface (Allen, 1987).
Physiologic factors affecting evaporative losses relate to an infant’s ability to sweat and to increase the minute ventilation. Although the physical characteristics of the newborn predispose him or her to heat loss, it has been demonstrated that neonates are capable of sweating in a warm environment (Bruck, 1961). Full-term neonates begin to sweat when rectal temperature reaches 37.5° to 37.9° C and ambient temperature exceeds 35° C. Although the onset of sweat production in infants who are small for gestational age is slower than it is in full-term infants, the maximum rates of sweat production are comparable (Sulyok et al., 1973). However, premature infants with a gestational age below 30 weeks show no sweating response because the lumen of their sweat glands are not yet fully developed.
Only a small amount of heat is lost when dry, inspired respiratory gases are humidified by water evaporating from the tracheobronchial epithelium. In adults, respiratory losses account for only 5% to 10% of total heat loss during anesthesia and surgery, and total insensible losses account for approximately 25% of the total heat dissipated (Bickler and Sessler, 1990). Minute ventilation on a per-kilogram basis in infants and children is significantly higher than in adults; thus, respiratory heat loss represents about one third of the total heat loss. Obviously, respiratory heat loss increases if the patient breathes cool, dry air as opposed to warm, moisturized air (Bissonnette et al., 1989a, 1989b).
Evaporative heat loss from a large surgical incision may equal all other sources of intraoperative heat loss combined (Roe, 1971). Because of increased evaporative heat loss, hypothermia is also more likely to occur if the skin of the patient is wet or comes in contact with wet drapes.
Heat generation
The ability to produce heat by increasing the metabolic rate and oxygen consumption is the other prerequisite of thermal regulation for a homeothermic organism (Hull and Smales, 1978). Beside the fact that three of the physical mechanisms leading to heat loss (i.e., conduction, radiation, and convection) can theoretically also be used to passively warm up a patient, the body has the ability to actively produce heat. Heat generation can be achieved through four mechanisms:
The behavioral aspect of heat production (voluntary muscle activity) is usually not functional in the perioperative period and therefore its role in heat production will not be discussed further here. Of the three remaining mechanisms for heat production, nonshivering thermogenesis is the major component in the newborn, whereas shivering thermogenesis is the main mechanism for heat production in older children and adults. The contribution of nonshivering thermogenesis in adults is debatable (Jessen, 1980a).
Although both the time course and the relationship between nonshivering and shivering thermogenesis in infants have been described, the exact time sequence and factors involved in the developmental aspects of switching on shivering thermogenesis and nonshivering thermogenesis off remain to be elucidated (Hull and Smales, 1978). The importance of nonshivering thermogenesis seems to decrease rapidly after the first year of life, while at the same time shivering thermogenesis is becoming more and more effective (Fig. 6-4B). Under normal conditions, dietary thermogenesis will not contribute significantly to thermogenesis during anesthesia. It only affects temperature if the patient is given food with a high protein or fructose content before or during anesthesia, which is normally not the case.
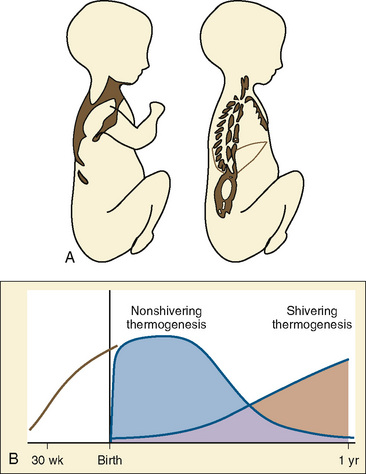
Nonshivering Thermogenesis
Brown fat differentiates in the human fetus between 20 and 30 weeks of gestational age (Lean et al., 1986a). It comprises only 2% to 6% of the infant’s total body weight and is found in six main locations: between the scapulae, in small masses around blood vessels of the neck, in large deposits in the axillae, in medium-size masses in the mediastinum, around the internal mammary vessels, and around the adrenal glands or kidneys (Fig. 6-4A).
Brown fat is a highly specialized tissue; the brown color is secondary to the abundant content of mitochondria in the cytoplasm of its multinucleated cells. These mitochondria are densely packed with cristae and have an increased content of respiratory-chain components (Himms-Hagen, 1976). They are unique in their ability to uncouple oxidative phosphorylation, resulting in heat production instead of generating adenosine triphosphate. This uncoupling process is mediated by the presence of the uncoupling protein 1 (UCP-1, or thermogenin) that is located on the inner mitochondrial membrane (Himms-Hagen, 1976; Ricquier and Kader, 1976).
Brown fat is highly vascularized and has a rich sympathetic innervation, which appears to be primarily β-sympathetic in origin and responsible for the uncoupling of oxidative phosphorylation (Karlberg et al., 1962, 1965). In respect to nonshivering thermogenesis, mature brown fat cells mainly rely on activation by β3-receptors. Cold stress increases sympathetic nervous system activity and norepinephrine release, which causes increased lipase activity in the brown fat tissue (Schiff et al., 1966). As a consequence, hydroxylation of triglycerides and release of free fatty acids occur. These free fatty acids act on UCP-1 and thereby increase the protein conductance across the mitochondrial membrane. In addition to norepinephrine, glucocorticoids and thyroxin have been implicated as factors that trigger nonshivering thermogenesis (Gale, 1973; Jessen, 1980b, 1980c). The heat produced by nonshivering thermogenesis is mainly a by-product of fatty acid metabolism, but to a minor degree it can also result from glucose metabolism. The activation of brown fat metabolism results in an increased proportion of the cardiac output being diverted through the brown fat. This proportion may reach as much as 25% of the cardiac output, which facilitates the direct warming of the blood.
Pharmacologic inhibition of nonshivering thermogenesis can be achieved with ganglionic and β-receptor blockade, inhalational anesthetics, and surgically by sympathectomy (Silverman et al., 1964; Stern et al., 1965; Ohlson et al., 1994). Inhibition of nonshivering thermogenesis by inhalational anesthetics starts as early as 5 minutes after turning on the vapor and starts to wean off within approximately 15 minutes after discontinuation of the inhalational anesthetic (Ohlson et al., 1994). Nonshivering thermogenesis is also inhibited in infants who have been anesthetized with fentanyl and propofol (Plattner et al., 1997).
In general, nonshivering thermogenesis seems to be quite variable in adults, but most often it does not appear to be functional or relevant (Ohlson et al., 1994; van Marken Lichtenbelt and Daanen, 2003). This assumption is supported by the fact that oxygen consumption does not increase significantly when patients exhibit thermoregulatory vasoconstriction (Mestyan et al., 1964; Dawkins and Scopes, 1965






Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


