The importance of nitric oxide (NO) in modulation of biological pathways has been known for over 20 years. The seminal studies by Furchgott and Zwadzki that provided an initial identification of a labile relaxing factor released from endothelial cells was a stimulus for many investigators to identify the nature of the transmitter and explore its role in biology (13). The tremendous volume of research papers on NO over the last two decades is a testament to the key physiologic and pathophysiologic roles that have been attributed to this molecule.
The wealth of knowledge surrounding this molecule is represented by a variety of excellent reviews that provide a detailed account of the molecular and cellular effects of this agent (see Forstermann et al. [12], Ignarro [22]). The aim of this manuscript is to provide a brief overview of how NO is generated by enzyme systems and how it can produce its biological actions with respect to its potential role in migraine.
NITRIC OXIDE: A KEY ROLE IN BIOLOGY
NO is a labile gas that has a short biological half-life, measured in seconds. It is a highly reactive and lipophilic molecule that has the ability to pass through biological membranes and interact with a number of target systems to produce its biological effects. The initial identification of NO as the endothelium-derived relaxing factor (13) produced by stimulating endothelial cells with agents such as acetylcholine or bradykinin focused attention on the role of NO in the cardiovascular system and control of blood pressure; however, there was a parallel development of our understanding of the potential role of NO in host defense (65) and as a neurotransmitter in the central nervous system (CNS) (55).
Perhaps one of the greatest initial breakthroughs came from the discovery that NO was produced by metabolism of the amino acid arginine by specific nitric oxide synthase (NOS) enzymes (4,5,22,34,55,65). Development of prototype tool inhibitor compounds that were simple analogs of arginine led to the use of these compounds to probe into the biological role of NO in physiology and pathophysiology. Many effects of NO were subsequently suggested and involved many organ systems and integrated biological pathways. NO is now believed to play a key role in many physiologic processes such as in the gastrointestinal tract, respiratory and cardiovascular systems, as well as the CNS. The first new medicines to modulate the NO pathway have also been developed for male erectile dysfunction.
CONTROL AND REGULATION OF NITRIC OXIDE PRODUCTION
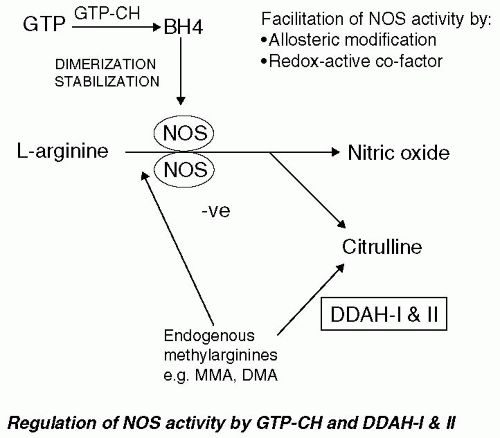
A schematic representation of the steps involved in the enzymatic production of NO is shown in Figure 15-1. It has been established that NO can be produced from a series of enzymes (NOS) from L-arginine with concomitant production of citrulline. Neuronal NOS was first isolated and characterized from rat brain and was found to have similarities with NADPH-cytochrome P450 reductase (4,5). Other isoforms of NOS were quickly discovered with the original nomenclature based on the site of purification with neuronal (nNOS), endothelial (eNOS) [39, XXX], and inducible (iNOS) (65) forms being located in macrophage. More recent studies have shown that NOS enzymes are widely expressed, are found in a range of cell types, and are not located specifically to the original location. An alternative nomenclature has been suggested with nNOS being NOS1, eNOS being NOS2, and iNOS being NOS3 (Table 15-1).
A number of critical cofactors have been described for NADPH, flavin adenine dincucleotide (FAD), and flavin adenine mononucleotide (FMN), heme, zinc (Zn), and tetrahydrobiopterin. FAD, FMN, Zn, and heme play key roles in the oxidation-reduction activity of NOS (34) and tetrahydrobiopterin plays a crucial role in dimerization of inactive NOS monomers and stabilization of the resultant functional enzyme (60).
FIGURE 15-1. Schematic representation of the steps involved in production of NO. L-Arginine is converted to NO and citrulline by the action of active NOS dimmers. Dimerization and stabilization of the active form occurs with tetrahydrobiopterin (BH4), which is synthesized from GTP by the action of GTP cyclohyrolase enzyme (GTP-CH). A number of critical cofactors have been described for NADPH, falvin adenine dincucleotide (FAD), and FMN, heme, Zn, and tetrahydrobiopterin. FAD, FMN, Zn, and heme play key roles in the oxidation-reduction activity of NOS. NOS activity is thought to be regulated by endogenous methylarginines, L-monomethyl arginine (L-NMMA), and ADMA. In normal circumstances the circulating concentration of L-NMMA and ADMA methylated arginines are maintained at low levels by the enzyme dimethylarginine dimethylaminohydrolase (DDAH I and II). (Adapted from Ignarro [3].)
Key properties of the NOS enzymes have been described in detail by other reviews (34,35) and are summarized in Table 15-1. It is striking that the NOS enzymes exhibit many similarities but with notable differences, particularly in the control of their expression and enzyme activity. For example, iNOS is not normally expressed but is under control of a variety of cellular stressors such as inflammatory cytokines and therefore is somewhat distinct from the other NOS isoforms. The distinct profile of iNOS in comparison to the other NOS isoforms is also supported by differences in sensitivity to intracellular calcium ion concentration. It is interesting to note that differences in the affinity of calmodulin, a calcium-binding protein, between iNOS and the other NOS isoforms are manifest in marked differences in the rate of NO production. Only small concentrations of NO are produced by the calcium-dependent NOS; NO production in these isozymes is reflected in the cellular calcium ion transient, whereas high concentrations can be produced by iNOS, as this enzyme activity is more related to transcriptional regulation.
Adapted from Forstermann et al. (2) and Ignarro (3).
Although transcriptional regulation of NOS activity was first identified in murine macrophage (65), it also appears possible for both nNOS and eNOS to be regulated transcriptionally, at least in some cell types (Table 15-1). Stressful stimuli such as hypoxia or increased glutamatergic activity have been shown to induce nNOS (8) and a variety of promoter sites have been identified that can control nNOS and eNOS expression. A key regulator for NOS expression appears to be nuclear factor κ B (NFκB). This appears to be especially important for the regulation of iNOS because stimuli as diverse as bacterial endotoxin, interleukins, and inflammatory cytokines have been shown to induce iNOS expression by translocation and activation of the NFκB pathway. NFκB is a transcription factor that provides a link between extracellular signaling and changes in gene expression. Following membrane receptor activation, the NFκB heterodimer translocates, through phosphorylation and degradation of inhibitory (IkB) subunits, to the nucleus where it binds to specific domains on DNA and induces gene expression.
NO can be formed in many cell types and is under close transcriptional and molecular control, for example, via regulation of intracellular calcium ion concentrations. Other means of physiologic regulation of the NO pathways are possible via production of endogenous inhibitors of NOS. For example, the NOS inhibitor L-NG-monomethyl arginine (L-NMMA) is a naturally occurring amino acid that is produced by protein arginine methyltransferases (62). Additionally, asymmetric dimethylargine (ADMA) is a nonselective inhibitor of NOS (32,33). The circulating concentration of L-NMMA and ADMA methylated arginines are maintained at low levels by the enzyme dimethylarginine dimethylaminohydrolase (DDAH) (Fig. 15-1), which catalyzes their conversion to citrulline, dimethylamine, or monomethylamine.
THE MOLECULAR TARGETS OF NITRIC OXIDE
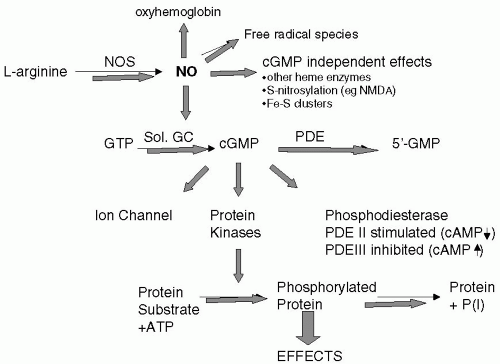
NO exhibits a range of unique features for a neurotransmitter or paracrine regulator with perhaps its most striking feature is the fact that it is a gaseous transmitter, highly reactive and unstable. NO does not interact with specific membrane-bound molecular targets in a similar manner as classical transmitters like acetylcholine at G-protein-coupled receptors or ion channels; NO acts via a series of redox reactions in a variety of proteins allowing marked diversity in the pathways linked to this molecule. These interactions may results in activation of biological pathways or inactivation of NO (Fig. 15-2). Perhaps the most well-characterized pathway is its binding to the heme group of guanylate cyclase resulting in activation of the enzyme and increased intracellular concentrations of cyclic guanosine monophosphate (cGMP), a key regulator of intracellular calcium ion concentrations and mediates cellular functions, such as vasodilatation. NO can therefore interact with heme-containing proteins. NO may also interact with non-heme-containing Fe and Fe-S-containing proteins as well as proteins with labile cysteine or tyrosine residues. NO can interact with a variety of other reactive oxygen or nitrogen species, which inactivate NO but can result in toxic derivatives such as peroxynitrite.
FIGURE 15-2. Schematic representation of potential fate of NO. NO may undergo a series of redox reactions that can inactivate the molecule and give rise to a diverse array of possible biological activities. NO may be inactivated by its binding to oxyhemoglobin or produce reactive peroxynitrite species. Activation of soluble guanylayte cyclase gives rise to a diverse array of possible effects in cells, by increasing the concentration of cGMP. cGMP levels in the cell are regulated by phosphodiesterase (PDE) enzymes that inactivate cGMP. cGMP may act directly to modulate channel function and other intracellular proteins phosphodiesterases. Activation of protein kinases produces phosphorylation of proteins and an array of biological effects. In addition, NO may bind directly to other heme-containing enzymes, nitrosylate other proteins (e.g., NMDA channel) or Fe-S-containing proteins. (Adapted from Ignarro [3].)
NO therefore has a potential multitude of effects and is involved in a number of key regulatory processes within the body. Some activities associated with NO are noteworthy in the consideration of the molecule as a mediator of migraine.
THE PHYSIOLOGIC EFFECTS OF NITRIC OXIDE
NO has a key role in regulation of cardiovascular tone and control of arterial blood pressure. The identification of endothelium-derived relaxing factor as NO, highlighted an autocrine role for this agent and implicated eNOS as a key source of NO in response to transmitters such as bradykinin or physiologic stimuli such as sheer stress. Work with genetically null mice in which the eNOS was deleted confirmed this role; there was marked hypertension and absence of acetylcholine-induced relaxation in isolated arteries (19). eNOS knockout animals are also more sensitive to cerebral ischemia (18) and have impaired cerebrovascular responses, highlighting the importance of eNOS-derived NO in control of physiologic processes.
NO clearly plays a modulatory role in a variety of systems and the complexity of these interactions is demonstrated by the neuroprotective effects of nitro-L-arginine in eNOS knockout animals (18). These indicated a detrimental role for non-eNOS-derived NO as the NOS inhibitor possessed neuroprotective effects in mice in which the beneficial effects of eNOS had been removed. These observations clearly support the concept of different NOS isoforms mediating opposing biological effects, in this case neuroprotection or neurotoxicity.
Within the CNS, NO plays a key role as a synaptic modulator and appears to play a central role in modulation of nociceptive pathways via a variety of mechanisms (see McMahon et al. [37] and Meller and Gebhart [38]). Studies show that NO can act as an retrograde transmitter facilitating glutamatergic transmission, possibly by the direct action of NO derivatives on the NR2B subunit of the N-methyl-D-aspartate (NMDA) channel (9) enhancing NMDA-evoked currents. NO plays a role in the modulation of pain pathways and learning and memory.
Only gold members can continue reading. Log In or Register to continue
 Principles of Clinical Pharmacology, Randomized Controlled Clinical Trials, and Evidence-Based Medicine in Headache
Principles of Clinical Pharmacology, Randomized Controlled Clinical Trials, and Evidence-Based Medicine in Headache
 Calcitonin Gene-Related Peptide and Other Peptides
Calcitonin Gene-Related Peptide and Other Peptides
 The Migraines: Introduction
The Migraines: Introduction
 Channelopathies and Their Possible Relation to Migraines
Channelopathies and Their Possible Relation to Migraines
 Autonomic Dysfunction in Migraines
Autonomic Dysfunction in Migraines
 Psychological and Behavioral Treatments of Migraines
Psychological and Behavioral Treatments of Migraines



