Calcitonin Gene-Related Peptide and Other Peptides
Calcitonin Gene-Related Peptide and Other Peptides
Susan Brain
Lars Edvinsson
Vasoactive peptides can be either stored or synthesized de novo before release from a range of tissues in the brain or from the walls of intracranial vasculature. In this chapter, we concentrate on neuropeptides that are released from perivascular nerves. These include calcitonin gene-related peptide (CGRP), substance P, neurokinin A, nociceptin, somatostatin, and opioids (Table 16-1). The endothelium produces the potent vasoconstrictors endothelin and angiotensin, and dilators such as nitric oxide, prostacyclin, and endothelium-derived hyperpolarizing factors. In addition there are circulating agents; among these the most potent is 5-hydroxytryptamine. The neuronal messengers stored in the intracranial vessels have been reviewed (32) and it was revealed that sympathetic nerves store noradrenaline, neuropeptide Y, and ATP, the parasympathetic fibers nitric oxide, acetylcholine, vasoactive intestinal peptide (VIP), peptide histidine isoleucine, pituitary adenylate cyclase-activating peptide (PACAP), and helodermine. Although VIP is of interest in certain forms of primary headaches, it is mainly the peptides contained in the sensory system, and this has attracted the most excitement during the last few years. A growing body of studies indicate that the distribution and biological activity of these neuropeptides to the areas of the cerebral vasculature and nervous system that are involved in migraine is highly relevant. Research into the precise role and importance of these peptides is often lacking because of the paucity of selective agents that either enhance or block activity or synthesis. However, there have been some major clinical trials that have focused on these peptides in recent years that have, for instance, ruled out a role for substance P, but provided evidence for the involvement of CGRP. This chapter is written in a comparative manner to allow an appreciation of our relative knowledge of the biology of these peptides and their role in migraine.
THE CGRP FAMILY OF PEPTIDES
The expression of mRNA from the calcitonin gene is tissue specific in that CGRP mRNA is predominantly expressed in nerves and calcitonin mRNA in the thyroid (5). The 37 amino acid peptide CGRP belongs to a family that include the more recently discovered peptides adrenomedullin that is primarily produced by non-neuronal tissues, especially vascular tissues and amylin that is mainly produced in the pancreas. They share some structural homology (approximately 25-40%) and also some, but not total, similarities in biological activities (see Brain and Grant [11] for recent review). CGRP is abundant in the body and has a wide distribution throughout the central and peripheral nervous systems. It has a number of biological activities, but the most relevant to migraine are its activities within the nervous and cardiovascular systems. CGRP is an extremely potent and long-lasting vasodilator, that is active at all levels of the cardiovascular system with good evidence for exquisite activity in the cerebral circulation (see Edvinsson [23]).
CGRP is most usually found in unmyelinated sensory C-fibers and myelinated Aδ-fibers that are commonly associated with blood vessels, where perivascular nerves terminate in close association with the vessels. Specific labeling of the C- and Aδ-fibers with wheat germ agglutinin-conjugated horse radish peroxidase and cholera toxin subunit b revealed that the C-fibers are located in lamina 1 and 2, and the Aδ-fibers from mechanoreceptors lay located in lamina 3 and 4 (48). Labeling experiments have revealed that the temporal artery and the superior sagittal sinus differ in their somatotopic organization. CGRP is the most prevalent of the neuropeptides in the sensory fibers. CGRP is commonly colocalized with other peptides in C-fibers, which include substance P (49). There are two forms of CGRP and αCGRP (or CGRPI), encoded by the calcitonin gene, that are relevant to the cerebral vasculature. βCGRP (or CGRPII), which has a high structural similarity (90%), is primarily found in the gut and formed from a distinct gene (57).
TABLE 16-1 Characteristics of Some Peptides Thought to be Involved in the Pathogenesis of Migraine
Neuropeptide
Amino Acids
Activity of Possible Relevance to Migraine
Location
Receptor Family
Modulates Animal Model of Migraine
Ligand Effective in Migraine
CGRP
37
Vasodilator
Sensory nerves
CGRP (CL/RAMP1)
Yes, antagonist (either CGRP8-37 or BIBN4096BS) decreases dural blood flow
NK1 receptor antagonists not effective in acute migraine
Somatostatin
14 (or 28)
Inhibits sensory nerve activity
Sensory nerves and other cells
Somatostatin (sst1-sst5)
Yes octa peptide benefits migraine pain
Opioids (enkephalins, etc.)
Varies
Inhibits sensory nerve activity
Sensory nerves and other cells
Opioid (μδκ and ORL-1)
Yes, inhibits dural plasma extravasation
Yes, but use limited by abusive nature
VIP
28
Vasodilator
Found in range of nerves
VIP (VPAC)
Not known
Not known
Neuropeptide Y
36
Vasoconstrictor
Sympathetic nerves
Y1-Y5
Not known
Not known
The distribution of CGRP-containing nerves has been evaluated in detail in the cerebral circulation (23). CGRP is contained in and released from sensory nerves originating in the trigeminal ganglia and innervates cerebral blood vessels (30). CGRP, when given intravenously, acts in a hypotensive manner, but it is generally considered that the major activity of CGRP is local to site of release (12). Plasma levels of CGRP are in the low picomolar level in normal volunteers (26) and migraineurs, but have been shown to increase in blood samples taken from the jugular vein ipsilateral to the attack (27).
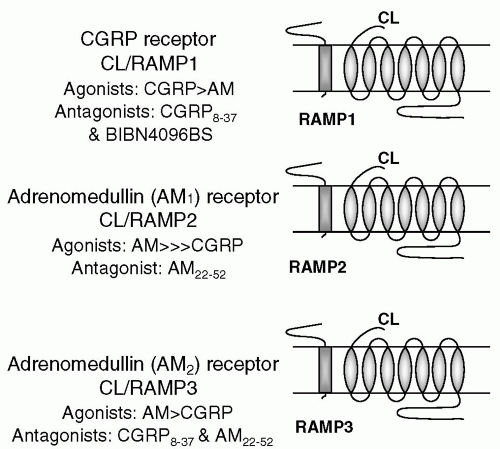
FIGURE 16-1. Summary of the characteristics of the CGRP receptor family.
The release of CGRP occurs in response to nerve stimulation, and this has been studied in tissues in response to chemical, physical, and mechanical stimulation in the laboratory. It is important to note that the TRPV (or VR-1) receptor for capsaicin, the hot extract of chili peppers, exists on the majority of CGRP-containing sensory nerves (13) both in animals and in human trigeminal ganglion (39). The endogenous stimuli for this receptor are under study and include protons, noxious heat, and a range of endogenous mediators, although their relative importance in pathology is unclear (2,3,4). Furthermore, presynaptic/prejunctional receptors on the sensory nerves can modulate CGRP release. These receptors include those for opioids, 5-hydroxytryptamine (5-HT1 receptor), γ-aminobutyric acid (GABAB receptor), histamine (H3 receptor), neuropeptide Y, somatostatin, VIP, purines, and galanin (see below and Maggi [51]).
CGRP receptors were classed as CGRP1 and CGRP2 in the late 1980s, as a consequence of pharmacologic studies. The CGRP1 receptor is considered the important cardiovascular receptor. The 8-37 amino acid fragment of CGRP, CGRP8-37, is a selective antagonist for this receptor (14). By comparison, the CGRP2 receptor remains poorly defined (60). It is now realized that the CGRP family of receptors consists of the seven transmembrane G protein-coupled calcitonin receptor-like receptor CL with one of three single membrane-spanning receptor activity modifying proteins (RAMPs; 53). This complex is associated with a CGRP-receptor component that is suggested to enhance receptor coupling and activation (25). The RAMP molecule is important for localization of the functional receptor to the cell surface and receptor phenotype, because it influences ligand specificity (65). CL is a G protein-coupled receptor that is important for ligand binding. Three RAMPs (RAMP1, RAMP2, and RAMP3) are known. CL, when presented as a heterodimer with RAMP1 at the cell surface, functions as a CGRP receptor that is antagonized by CGRP8-37 and the nonpeptide CGRP antagonist BIBN4096BS (see below). CL with RAMP2 produces an adrenomedullin (AM) receptor that is blocked by the weak AM antagonist ADM22-52. CL with RAMP3 leads to an AM receptor that least is known about. Both CL and the RAMPs have been detected in the human cerebral and meningeal vasculature (59).
The best described, and only potent, small molecule CGRP antagonist described to date has been BIBN4096BS, 1-Piperidinecarboxamide, N-[2-[[5-amino-1-[[4-(4-pyridinyl)-1-piperazinyl]carbonyl] pentyl] amino]-1-[(3,5-dibromo-4-hydroxyphenyl)methyl]-2-oxoethyl]-4-(1,4-dihydro-2-oxo-3(2H)-quinazolinyl)-,[R-(R*,S*)]. It is a competitive antagonist with potent properties at the human and marmoset CGRP1 receptor (18). It displays a 200-fold greater affinity in human compared with rodent tissues. RAMP1 governs species selectivity for receptor antagonists, via a single amino acid residue (tryptophan at position 74; see Mallee et al. [52]). A related compound, compound 1, (4-(2-Oxo-2, 3-dihydro-benzoimidazol-1-yl)-piperidine-1-carboxylic acid [1-3,5-dibromo-4-hydroxy-benzyl)-2-oxo-2-(4-phenyl-piperazin-1-yl)-ethyl]-amide) has also been synthesized and studied. It is a weak antagonist of CGRP receptors. It also weakly antagonized CGRP responses in human cerebral and guinea pig basilar arteries (22), but failed to inhibit the vascular relaxation induced by CGRP in porcine tissues (33), where CGRP8-37 is an effective antagonist. However, compound 1 acted as a competitive antagonist at the CGRP receptor in human left anterior descending coronary arteries having a pA2 value similar to that of CGRP8-37 (34). This emphasizes the species and tissue selectivity that is now becoming associated with CGRP receptor antagonists. Alternatively, a distinct nonpeptide CGRP1 receptor antagonist that acts across species exists. SB-273779, [N-methyl-N-(2-methylphenyl)-3-nitro-4-(2-thiazolylsulfinyl)nitrobenzanilide] is selective for the CGRP receptor. The compound is weaker than BIBN4096BS, but does displace radioactive CGRP in rat and porcine lung binding studies (1).
Only gold members can continue reading. Log In or Register to continue
 Principles of Clinical Pharmacology, Randomized Controlled Clinical Trials, and Evidence-Based Medicine in Headache
Principles of Clinical Pharmacology, Randomized Controlled Clinical Trials, and Evidence-Based Medicine in Headache
 Nitric Oxide
Nitric Oxide
 The Migraines: Introduction
The Migraines: Introduction
 Channelopathies and Their Possible Relation to Migraines
Channelopathies and Their Possible Relation to Migraines
 Autonomic Dysfunction in Migraines
Autonomic Dysfunction in Migraines
 Psychological and Behavioral Treatments of Migraines
Psychological and Behavioral Treatments of Migraines



